Гормональная регуляция обмена веществ
Гормон роста оказывает мощное липолитическое действие, которое отличается от действия катехоламинов. Он вызывает увеличение концентрации свободных жирных кислот в плазме через 2-3 часа. Это действие, отмечающееся при введении даже небольших доз СТГ, по-видимому, связано с торможением процесса реэстерификации свободных жирных кислот. Тем не менее СТГ оказывает определенное модулирующее влияние и на активность аденилатциклазы.
Другие гипофизарные гормоны (АКТГ, ТТГ, меланоцитостимулирующий гормон) также оказывают липолитическое действие, хотя и менее выраженное, чем у СТГ. Большинство авторов считают, что оно осуществляется путем активации аденилатциклазы и повышения скорости образования цАМФ.
Тироидные гормоны и кортикостероиды оказывают пермиссивное (разрешающее) действие на липолиз, поскольку липолитический и калоригенный эффекты катехоламинов не проявляются в отсутствие кортикостероидов и тироидных гормонов. Имеются данные о том, что ТТГ, АКТГ, гормоны щитовидной железы и коры надпочечников индуцируют синтез аденилатциклазы и, следовательно, принимают непосредственное участие в стимуляции липолиза. Кроме того, тироидные гормоны влияют на активность ацил-КоА-синтетазы, локализующейся в митохондриях, и цитоплазматического фермента глицеролфосфатдегидрогеназы. Эти ферменты участвуют в регуляции скорости липолиза и липогенеза.
Как уже отмечалось, инсулин обладает характерным антилиполитическим свойством, и при сахарном диабете вследствие увеличения липолиза повышается концентрация свободных жирных кислот в плазме, снижению которой способствует инсулинотерапия.
Белковый обмен. Белки в организме также являются источником энергии. Они содержатся главным образом в мышцах и их количество составляет в организме здорового человека массой 70 кг около 6000 г, что соответствует 24 000 ккал. Циркуляция их в крови в виде аминокислот незначительна и составляет всего 6 г, или 24 ккал. Белки – необходимый компонент любой ткани организма – поступают в организм с пищей и в желудочно-кишечном тракте после воздействия на них ферментов (пепсина, трипсина) гидролизуются до небольших пептидов и аминокислот, которые затем всасываются в кровь и лимфу. В организме человека для синтеза пуринов, пиримидинов, порфиринов используются только аминокислоты, поэтому все поступающие с пищей белки должны быть диссоциированы в различных ферментативных реакциях до отдельных аминокислот.
Некоторые аминокислоты могут синтезироваться в организме, поэтому называются заменимыми: аланин, аспарагиновая кислота, цистеин, глутаминовая кислота, глицин, пролин, серин, тирозин, аспарагин, глутамин; другие же не могут быть синтезированы и называются незаменимыми: лейцин, изолейцин, валин, лизин, метионин, фенилаланин,
триптофан, треонин, гистидин, аргинин (гистидин и аргинин синтезируются в организме взрослого человека).Синтез белка – сложный процесс, происходящий постоянно. Информация о структуре любого белка данного организма хранится в хромосомах в виде генетического кода. При поступлении сигнала о необходимости синтеза определенного белка с участка ДНК, на котором закодирована структура данного белка, при участии фермента РНК-полимеразы начинает образовываться мРНК. Процесс образования мРНК называется “транскрипция”. Если молекула ДНК относительно стабильна, то период полураспада мРНК составляет 2-80 ч (время, необходимое для синтеза белка).
Образовавшаяся мРНК покидает ядро и направляется к рибосомам, где и осуществляется синтез белка. На рибосомах локализуются рибосомальная РНК (рРНК) и транспортная РНК (тРНК), которые вместе участвуют в процессе считывания информации, заложенной в мРНК, и“сборки” нового белка. Обычно рРНК и метионил-тРНК присоединяются к специальной точке мРНК, и с этого момента начинается их движение вдоль молекулы мРНК, во время которого “считываются” триплетные кодоны и начинается “сборка” полипептидной цепи нового белка.
Аминокислоты могут использоваться рибосомами лишь после их активации соответствующими ферментами, число которых по всей вероятности соответствует количеству аминокислот.
Гормоны влияют на скорость синтеза белка несколькими путями: повышением или снижением активности ферментов, участвующих в процессе “сборки” белковой молекулы, изменением транспорта аминокислот, необходимых для синтеза белка, повышением активности и скорости образования рибосом, осуществляющих синтез белка, повышением скорости начала образования полисом, повышением активности РНК-полимеразы и скорости образования мРНК. Некоторые вещества могут в той или иной степени ингибировать перечисленные процессы. Так, актиномицин, связываясь с ДНК, угнетает синтез мРНК и таким образом препятствует действию гормонов на процессы транскрипции. Пуромицин угнетает синтез белка, влияя на тРНК, и ингибирует гормональную регуляцию процессов трансляции.
Наряду с процессами синтеза белка в организме постоянно протекают процессы белкового катаболизма, конечным продуктом которого у человека является мочевина. Аммиак, образующийся в большинстве клеток в процессе катаболизма аминокислот, наряду с другими азотсодержащими веществами превращается в печени в мочевину в цикле Кребса-Гензелейта (схема 10). При этом аммиак транспортируется в печень не в свободной форме, а детоксицируется глутаматом с образованием глутамина под влиянием фермента глутаминсинтетазы. В митохондриях печени глутамин при участии глутаминазы вновь диссоциирует на аммиак и глутамат. Последний возвращается в ткани, а аммиак используется для образования карбамоилфосфата с помощью фермента карбамоилфосфатсинтетазы.
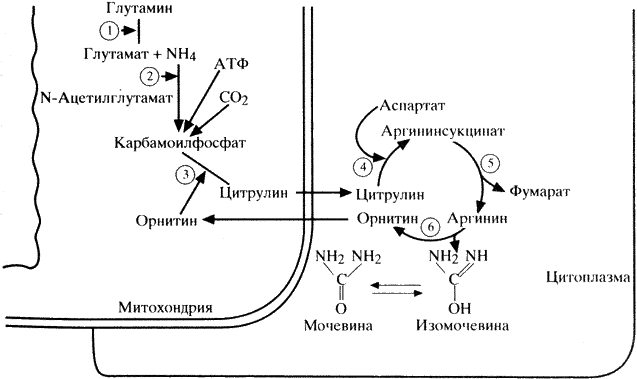
1 – глутаминаза; 2 – карбамоилфосфатсинтетаза; 3 – орнитинкарбамоилтрансфераза; 4 – аргининсукци- натсинтетаза; 5 – аргининсукцинатлиаза; 6 – аргиназа.
Другим основным для начального биосинтеза мочевины митохондриальным ферментом является орнитинкарбамоилтрансфераза, катализирующая образование цитрулина из карбамоилфосфата и орнитина. Цитрулин через митохондриальную мембрану проникает в цитоплазму клетки, где взаимодействует с аспартатом при участии аргиносукцинатсинтетазы, в результате чего образуется аргиносукцинат, который под воздействием аргиносукцинатлиазы диссоциирует на аргинин и фурамат. Аргинин с помощью аргиназы распадается на мочевину и орнитин, транспортируемый обратно в митохондрии, где он вступает в новый цикл образования цитрулина. Для образования одной молекулы мочевины в цикле Кребса-Гензелейта используется 4 молекулы АТФ (Бонди Ф., Розенберг Л., 1980).
В зависимости от путей катаболизма различают глюкогенные, кетогенные и смешанные аминокислоты. Кетогенной аминокислотой является лейцин, который распадается на ацетоуксусную кислоту и ацетил-КоА, вызывающие повышение уровня кетоновых тел в крови. Изолейцин, лизин, фенилаланин и тирозин – глюкогенные и кетогенные аминокислоты. Фенилаланин и тирозин распадаются на фурамат и ацетоацетат, которые могут быть использованы в процессах глюконеогенеза. К глюкогенным аминокислотам относятся аланин, аргинин, аспарагиновая кислота, цистеин, глутаминовая кислота, глицин, гистидин, гидроксипролин, метионин, серин, треонин, триптофан, валин. Продукты распада этих аминокислот участвуют в процессах глюконеогенеза.
Количество аминокислот в сыворотке крови поддерживается постоянно на определенном уровне за счет поступления их из желудочно-кишечного тракта и депо, которыми являются печень и мышцы. В мышцах содержится более 50% общего количества свободных аминокислот организма. Наиболее мобильны из них аланин и глутамин, составляющие более 50% всех аминокислот, высвобождающихся из мышц. Аланин синтезируется в мышцах путем трансаминирования пирувата. Глутамин поступает в почки, где отщепляющийся азот используется для образования аммиака. Аланин же задерживается печенью, где быстро конвертируется в глюкозу через образование пирувата. Последний процесс получил название цикла аланина и наряду с циклом лактата (цикл Кори) имеет большое значение в процессах глюконеогенеза. Взаимоотношения этих двух циклов представлены на схеме 11.
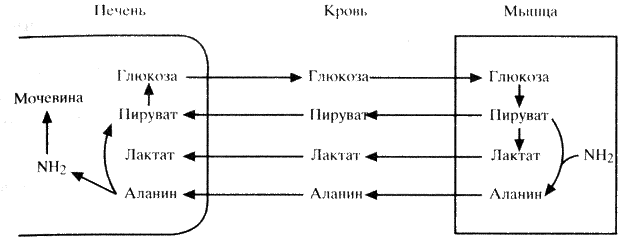
Эндокринная система участвует в регуляции белкового обмена как непосредственно, так и опосредованно – путем первичного влияния на углеводный, жировой и минеральный обмен.
По воздействию на белковый обмен все гормоны можно разделить на две большие группы: 1) гормоны, первичное действие которых – стимуляция процессов транскрипции – проявляется в ядре клетки (стероидные и тироидные гормоны); 2) гормоны, стимулирующие процессы трансляции, происходящие дистальнее ядра клетки.
Анаболическое действие инсулина заключается в ускорении проникновения аминокислот через мембрану клетки и включения их в белки, что вызывает снижение уровня аминокислот в крови. Это действие инсулина не зависит от синтеза РНК и не угнетается актиномицином. Инсулин снижает активность аминотрансфераз и ферментов цикла мочевины. Последний эффект инсулина характеризуется повышением активности РНК-полимеразы и концентрации РНК в печени. При этом увеличивается скорость образования полисом и рибосом.
Действие инсулина на синтез белка в печени противоположно действию глюкагона, который усиливает катаболизм аминокислот и мобилизует их для процессов глюконеогенеза.
Основным эффектом действия гормона роста является стимуляция анаболичесих процессов, чем и обусловливается ускорение роста скелета при гиперсекреции этого гормона. Он, так же как инсулин, стимулирует транспорт аминокислот и их включение рибосомами в белки, повышает скорость образования полисом и рибосом, активность РНК-полимеразы, увеличивает количество РНК в клетках, что сопровождается повышением скорости синтеза белков. Гормон роста угнетает активность ферментов, разрушающих аминокислоты, повышает уровень инсулиноподобных факторов роста I и II (ИФР I и II) в сыворотке крови, стимулирует синтез коллагена в костях, коже, других органах и тканях организма.
Введение в кровь тироидных гормонов сопровождается усилением основного обмена, тахикардией, уменьшением содержания гликогена в мышце сердца, увеличением чувствительности тканей к липолитическому действию адреналина. Влияние тироидных гормонов на обмен веществ обнаруживается лишь через 48 часов и более, т.е. значительно позже, чем действие инсулина и СТГ.
Усиление основного обмена после введения тироидных гормонов является следствием увеличения количества и размера митохондрий, количества рибосом, содержания в них цитохрома и образования митохондриальной мембраны. Тироидные гормоны не влияют на транспорт аминокислот. Под действием этих гормонов возрастает активность ферментов, в том числе глицеролфосфатдегидрогеназы, цитохромоксидазы, цитохрома С, NAD-изолимонной дегидрогеназы, а также РНК-полимеразы и увеличивается синтез РНК (включая мРНК).
Под влиянием эстрогенов увеличивается содержание РНК, особенно в тканях-мишенях, и количество белка, повышаются активность РНК-полимеразы и скорость синтеза РНК. Считается, что после связывания эстрогенов с негистоновыми белками хроматина ядра происходит стимуляция определенной области генома, образование новых видов РНК и некоторых специфических (кислых) белков. Длительность первой фазы действия эстрогенов от нескольких минут до 1-2 ч. После этого вновь образовавшиеся белки увеличивают содержание РНК, что приводит к усилению синтеза белка. Длительность второй фазы гормонального эффекта составляет 4 ч и более.
Андрогены оказывают на белковый обмен такое же действие, как и эстрогены. Кроме того, тестостерон и другие андрогены влияют на развитие мышечной ткани, улучшая транспорт аминокислот и повышая активность РНК-полимеразы.
При введении больших доз глюкокортикоидов наблюдается быстрое уменьшение мышечной массы и снижение скорости включения аминокислот в белки мышц, селезенки, костной ткани, отмечается снижение активности РНК-полимеразы и способности изолированных рибосом поглощать (включать) аминокислоты. Эти изменения не распространяются на печень, где увеличивается синтез ферментов, участвующих в процессах глюконеогенеза и разрушения аминокислот, в том числе глюкозо-6-фосфатазы, фосфоенолпируваткарбоксикиназы, пируваткарбоксилазы и др.
Таким образом, гормоны регулируют все виды обмена веществ (углеводный, жировой, белковый, минеральный и др.), поддерживая постоянство внутренней среды, которое, как установил еще Клод Бернар, является непременным условием жизнедеятельности организма.

Ваш комментарий