Углеводный обмен
Углеводный обмен — совокупность процессов превращения моносахаридов и их производных, а также гомополисахаридов, гетерополисахаридов и различных углеводсодержащих биополимеров (гликоконъюгатов) в организме человека и животных. В результате У. о. происходит снабжение организма энергией (см. Обмен веществ и энергии), осуществляются процессы передачи биологической информации и межмолекулярные взаимодействия, обеспечиваются резервные, структурные, защитные и другие функции углеводов. Углеводные компоненты многих веществ, например гормонов, ферментов, транспортных гликопротеинов, являются маркерами этих веществ, благодаря которым их «узнают» специфические рецепторы плазматических и внутриклеточных мембран.
Синтез и превращения глюкозы в организме. Один из наиболее важных углеводов — глюкоза — является не только основным источником энергии, но и предшественником пентоз, уроновых кислот и фосфорных эфиров гексоз. Глюкоза образуется из гликогена и углеводов пищи — сахарозы, лактозы, крахмала, декстринов. Кроме того, глюкоза синтезируется в организме из различных неуглеводных предшественников (рис. 1). Этот процесс носит название глюконеогенеза и играет важную роль в поддержании гомеостаза. В процессе глюконеогенеза участвует множество ферментов и ферментных систем, локализованных в различных клеточных органеллах. Глюконеогенез происходит главным образом в печени и почках.
Существуют два пути расщепления глюкозы в организме: гликолиз (фосфоролитический путь, путь Эмбдена — Мейергофа — Парнаса) и пентозофосфатный путь (пентозный путь, гексозомонофосфатный шунт). Схематически пентозофосфатный путь выглядит так: глюкозо-6-фосфат ® 6-фосфатглюконолактон ® рибулозо-5-фосфат ® рибозо-5-фосфат. В ходе пентозофосфатного пути происходит последовательное отщепление от углеродной цепи сахара по одному атому углерода в виде СО2. В то время как гликолиз играет важную роль не только в энергетическом обмене, но и в образовании промежуточных продуктов синтеза липидов, пентозофосфатный путь приводит к образованию рибозы и дезоксирибозы, необходимых для синтеза нуклеиновых кислот (ряда коферментов.
Синтез и распад гликогена. В синтезе гликогена — главного резервного полисахарида человека и высших животных — участвуют два фермента: гликогенсинтетаза (уридиндифосфат (УДФ) глюкоза: гликоген-4a-глюкозилтрансфераза), катализирующая образование полисахаридных цепей, и ветвящий фермент, образующий в молекулах гликогена так называемые связи ветвлении. Для синтеза гликогена необходимы так называемые затравки. Их роль могут выполнять либо глюкозиды с различной степенью полимеризации, либо белковые предшественники, к которым при участии особого фермента глюкопротеинсинтетазы присоединяются глюкозные остатки уридиндифосфатглюкозы (УДФ-глюкозы).
Распад гликогена осуществляется фосфоролитическим (гликогенолиз) или гидролитическим путями. Гликогенолиз представляет собой каскадный процесс, в котором участвует ряд ферментов фосфорилазной системы — протеинкиназа, киназа фосфорилазы b, фосфорилаза b, фосфорилаза а, амило-1,6-глюкозидаза, глюкозо-6-фосфатаза. В печени в результате гликогенолиза образуется глюкоза из глюкозо-6-фосфата благодаря действию на него глюкозо-6-фосфатазы, отсутствующей в мышцах, где превращения глюкозо-6-фосфата приводят к образованию молочной кислоты (лактата). Гидролитический (амилолитический) распад гликогена (рис. 2) обусловлен действием ряда ферментов, называемых амилазами (a-глюкозидазами). Известны a-, b- и g-амилазы. a-Глюкозидазы в зависимости от локализации в клетке делят на кислые (лизосомные) и нейтральные.
Синтез и распад углеводсодержащих соединений. Синтез сложных сахаров и их производных происходит с помощью специфических гликозилтрансфераз, катализирующих перенос моносахаридов от доноров — различных гликозилнуклеотидов или липидных переносчиков к субстратам-акцепторам, которыми могут быть углеводный остаток, полипептид или липид в зависимости от специфичности трансфераз. Нуклеотидным остатком является обычно дифосфонуклеозид.
В организме человека и животных много ферментов, ответственных за превращение одних углеводов в другие, как в процессах гликолиза и глюконеогенеза, так и в отдельных звеньях пентозофосфатного пути.
Ферментативное расщепление углеводсодержащих соединений происходит в основном гидролитическим путем с помощью гликозидаз, отщепляющих углеводные остатки (экзогликозидазы) или олигосахаридные фрагменты (эндогликозидазы) от соответствующих гликоконъюгатов. Гликозидазы являются чрезвычайно специфическими ферментами. В зависимости от природы моносахарида, конфигурации его молекулы (их D или L-изомеров) и типа гидролизуемой связи (a или b) различают a—D-маннозидазы, a—L-фукозидазы, ×b—D-галактозидазы и т.д. Гликозидазы локализованы в различных клеточных органеллах; многие из них локализованы в лизосомах. Лизосомные (кислые) гликозидазы отличаются от нейтральных не только локализацией в клетках, оптимальным для их действия значением рН и молекулярной массой, но и электрофоретической подвижностью и рядом других физико-химических свойств.
Гликозидазы играют важную роль в различных биологических процессах; они могут, например, оказывать влияние на специфический рост трансформированных клеток, на взаимодействие клеток с вирусами и др.
Имеются данные о возможности неферментативного гликозилирования белков in vivo, например гемоглобина, белков хрусталика, коллагена. Есть сведения, что неферментативное гликозилирование (гликирование) играет важную патогенетическую роль при некоторых заболеваниях (сахарном диабете, галактоземии и др.).
Транспорт углеводов. Переваривание углеводов начинается в ротовой полости при участии гидролитических ферментов слюны. Гидролиз ферментами слюны продолжается в желудке (сбраживание углеводов пищевого комка предотвращается соляной кислотой желудочного сока). В двенадцатиперстной кишке полисахариды пищи (крахмал, гликоген и др.) и сахара (олиго- и дисахариды) расщепляются при участии a-глюкозидаз и других гликозидаз сока поджелудочной железы до моносахаридов, которые всасываются в тонкой кишке в кровь. Скорость всасывания углеводов различна, быстрее всасываются глюкоза и галактоза, медленнее — фруктоза, манноза и другие сахара.
Транспорт углеводов через эпителиальные клетки кишечника и поступление в клетки периферических тканей осуществляются с помощью особых транспортных систем, функция которых заключается и переносе молекул сахаров через клеточные мембраны. Существуют особые белки-переносчики — пермеазы (транслоказы), специфические по отношению к сахарам и их производным. Транспорт углеводов может быть пассивным и активным. При пассивном транспорте перенос углеводов осуществляется по направлению градиента концентрации, так что равновесие достигается тогда, когда концентрации сахара в межклеточном веществе или межклеточной жидкости и внутри клеток выравниваются. Пассивный транспорт сахаров характерен для эритроцитов человека. При активном транспорте углеводы могут накапливаться в клетках и концентрация их внутри клеток становится выше, чем в окружающей клетки жидкости. Предполагают, что активное поглощение сахаров клетками отличается от пассивного тем, что последнее является Na+-независимым процессом. В организме человека и животных активный транспорт углеводов происходит главным образом в клетках эпителия слизистой оболочки кишечника и в извитых канальцах (проксимальных отделах нефрона) почек.
Регуляция углеводного обмена осуществляется при участии очень сложных механизмов, которые могут оказывать влияние на индуцирование или подавление синтеза различных ферментов У. о. либо способствовать активации или торможению их действия. Инсулин, катехоламины, глюкагон, соматотропный и стероидные гормоны оказывают различное, но очень выраженное влияние на разные процессы углеводного обмена. Так, например, инсулин способствует накоплению в печени и мышцах гликогена, активируя фермент гликогенсинтетазу, и подавляет гликогенолиз и глюконеогенез. Антагонист инсулина — глюкагон стимулирует гликогенолиз. Адреналин, стимулируя действие аденилатциклазы, оказывает влияние на весь каскад реакций фосфоролиза. Гонадотропные гормоны активируют гликогенолиз в плаценте. Глюкокортикоидные гормоны стимулируют процесс глюконеогенеза. Соматотропный гормон оказывает влияние на активность ферментов пентозофосфатного пути и снижает утилизацию глюкозы периферическими тканями. В регуляции глюконеогенеза принимают участие ацетил-КоА и восстановленный никотинамидадениндинуклеотид. Повышение содержания жирных кислот в плазме крови тормозит активность ключевых ферментов гликолиза. В регуляции ферментативных реакций У. о. важную цель играют ионы Са2+, непосредственно или при участии гормонов, часто в связи с особым Са2+-связывающим белком — калмодулином. В регуляции активности многих ферментов большое значение имеют процессы их фосфорилирования — дефосфорилирования. В организме существует прямая связь между У. о. и обменом белков (см. Азотистый обмен), липидов (см. Жировой обмен) и минеральных веществ (см. Минеральный обмен).
Патология углеводного обмена. Увеличение содержания глюкозы в крови — гипергликемия может происходить вследствие чрезмерно интенсивного глюконеогенеза либо в результате понижения способности утилизации глюкозы тканями, например при нарушении процессов ее транспорта через клеточные мембраны. Понижение содержания глюкозы в крови — гипогликемия — может являться симптомом различных болезней и патологических состояний, причем особенно уязвимым в этом отношении является мозг: следствием гипогликемии могут быть необратимые нарушения его функций.
Генетически обусловленные дефекты ферментов У. о. являются причиной многих наследственных болезней. Примером генетически обусловленного наследственного нарушения обмена моносахаридов может служить галактоземия, развивающаяся в результате дефекта синтеза фермента галактозо-1-фосфатуридилтрансферазы. Признаки галактоземии отмечают также при генетическом дефекте УДФ-глюкоза-4-эпимеразы. Характерными признаками галактоземии являются гипогликемия, галактозурия, появление и накопление в крови наряду с галактозой галактозо-1-фосфата, а также снижение массы тела, жировая дистрофия и цирроз печени, желтуха, катаракта, развивающаяся в раннем возрасте, задержка психомоторного развития. При тяжелой форме галактоземии дети часто погибают ни первом году жизни вследствие нарушений функций печени или пониженной сопротивляемости инфекциям.
Примером наследственной непереносимости моносахаридов является непереносимость фруктозы, которая вызывается генетическим дефектом фруктозофосфатальдолазы и в ряде случаев — снижением активности Фруктоза-1,6-дифосфат-альдолазы. Болезнь характеризуется поражениями печени и почек. Для клинической картины характерны судороги, частая рвота, иногда коматозное состояние. Симптомы заболевания появляются в первые месяцы жизни при переводе детей на смешанное или искусственное питание. Нагрузка фруктозой вызывает резкую гипогликемию.
Заболевания, вызванные дефектами в обмене олигосахаридов, в основном заключаются в нарушении расщепления и всасывания углеводов пищи, что происходит главным образом в тонкой кишке. Мальтоза и низкомолекулярные декстрины, образовавшиеся из крахмала и гликогена пищи под действием a-амилазы слюны и сока поджелудочной железы, лактоза молока и сахароза расщепляются дисахаридазами (мальтазой, лактазой и сахаразой) до соответствующих моносахаридов в основном в микроворсинках слизистой оболочки тонкой кишки, а затем, если процесс транспорта моносахаридов не нарушен, происходит их всасывание. Отсутствие или снижение активности дисахаридаз к слизистой оболочке тонкой кишки служит главной причиной непереносимости соответствующих дисахаридов, что часто приводит к поражению печени и почек, является причиной диареи, метеоризма (см. Мальабсорбции синдром). Особенно тяжелыми симптомами характеризуется наследственная непереносимость лактозы, обнаруживающаяся обычно с самого рождения ребенка. Для диагностики непереносимости сахаров применяют обычно нагрузочные пробы с введением натощак per os углевода, непереносимость которого подозревают. Более точный диагноз может быть поставлен путем биопсии слизистой оболочки кишечника и определения в полученном материале активности дисахаридаз. Лечение состоит в исключении из пищи продуктов, содержащих соответствующий дисахарид. Больший эффект наблюдают, однако, при назначении ферментных препаратов, что позволяет таким больным употреблять обычную пищу. Например, в случае недостаточности лактазы, содержащий ее ферментный препарат, желательно добавлять в молоко перед употреблением его в пищу. Правильный диагноз заболеваний, вызванных недостаточностью дисахаридаз, крайне важен. Наиболее частой диагностической ошибкой в этих случаях являются установление ложного диагноза дизентерии, других кишечных инфекций, и лечение антибиотиками, приводящее к быстрому ухудшению состояния больных детей и тяжелым последствиям.
Заболевания, вызванные нарушением обмена гликогена, составляют группу наследственных энзимопатий, объединенных под названием гликогенозов. Гликогенозы характеризуются избыточным накоплением гликогена в клетках, которое может также сопровождаться изменением структуры молекул этого полисахарида. Гликогенозы относят к так называемым болезням накопления. Гликогенозы (гликогенная болезнь) наследуются по аутосомно-рецессивному или сцепленному с полом типу. Почти полное отсутствие в клетках гликогена отмечают при агликогенозе, причиной которого является полное отсутствие или сниженная активность гликогенсинтетазы печени.
Заболевания, вызванные нарушением обмена различных гликоконъюгатов, в большинстве случаев являются следствием врожденных нарушений распада гликолипидов, гликопротеинов или гликозаминогликанов (мукополисахаридов) в различных органах. Они также являются болезнями накопления. В зависимости от того, какое соединение аномально накапливается в организме, различают гликолипидозы, гликопротеиноды, мукополисахаридозы. Многие лизосомные гликозидазы, дефект которых лежит в основе наследственных нарушений углеводного обмена, существуют в виде различных форм,
так называемых множественных форм, или изоферментов. Заболевание может быть вызвано дефектом какого-либо одного изофермента. Так, например. болезнь Тея — Сакса — следствие дефекта формы AN-ацетилгексозаминидазы (гексозаминидазы А), в то время как дефект форм А и В этого фермента приводит к болезни Сандхоффа.Большинство болезней накопления протекает крайне тяжело, многие из них пока неизлечимы. Клиническая картина при различных болезнях накопления может быть сходной, и, напротив, одно и то же заболевание может проявляться по-разному у разных больных. Поэтому необходимо в каждом случае устанавливать ферментный дефект, выявляемый большей частью в лейкоцитах и фибробластах кожи больных. В качестве субстратов применяют гликоконьюгаты или различные синтетические гликозиды. При различных мукополисахаридозах, а также при некоторых других болезнях накопления (например, при маннозидозе) выводятся с мочой в значительных количествах различающиеся по структуре олигосахариды. Выделение этих соединений из мочи и их идентификацию проводят с целью диагностики болезней накопления. Определение активности фермента в культивируемых клетках, выделенных из амниотической жидкости, получаемой при амниоцентезе при подозрении на болезнь накопления, позволяет ставить пренатальный диагноз.
При некоторых заболеваниях серьезные нарушения У. о. возникают вторично. Примером такого заболевания является диабет сахарный, обусловленный либо поражением b-клеток островков поджелудочной железы, либо дефектами в структуре самого инсулина или его рецепторов на мембранах клеток инсулинчувствительных тканей. Алиментарные гипергликемия и гиперинсулинемия ведут к развитию ожирения, что увеличивает липолиз и использование неэтерифицированных жирных кислот (НЭЖК) в качестве энергетического субстрата. Это ухудшает утилизацию глюкозы в мышечной ткани и стимулирует глюконеогенез. В свою очередь, избыток в крови НЭЖК и инсулина ведет к увеличению синтеза в печени триглицеридов (см. Жиры) и холестерины и, соответственно, к увеличению концентрации в крови липопротеинов очень низкой и низкой плотности. Одной из причин, способствующих развитию таких тяжелых осложнений при диабете, как катаракта, нефропатия, англопатия и гипоксия тканей, является неферментативное гликозилирование белков.
Особенности углеводного обмена у детей. Состояние У. о. у детей в норме определяется зрелостью эндокринных механизмов регуляции и функций других систем и органов. В поддержании гомеостаза плода важную роль играет поступление к нему глюкозы через плаценту. Количество глюкозы, поступающей через плаценту к плоду, непостоянно, т.к. ее концентрация в крови матери может неоднократно меняться в течение дня. Изменение соотношения инсулин/глюкоза у плода может вызвать у него острые или длительные нарушения обмена веществ. В последнюю треть внутриутробного периода у плода значительно увеличиваются запасы гликогена в печени и мышцах, в этот период глюкогенолиз и глюконеогенез уже имеют для плода существенное значение и как источник глюкозы.
Особенностью У. о. у плода и новорожденного является высокая активность процессов гликолиза, позволяющая лучше адаптироваться к условиям гипоксии. Интенсивность гликолиза у новорожденных на 30—35% выше, чем у взрослых; в первые месяцы после рождения она постепенно снижается. О высокой интенсивности гликолиза у новорожденных свидетельствуют высокое содержание лактата в крови и моче и более высокая, чем у взрослых, активность лактатдегидрогеназы в крови. Значительная часть глюкозы у плода окисляется по пентозофосфатному пути.
Родовой стресс, изменение температуры окружающей среды, появление самостоятельного дыхания у новорожденных, возрастание мышечной активности и усиление деятельности мозга увеличивают расход энергии во время родов и в первые дни жизни, приводя к быстрому снижению содержания глюкозы в крови. Через 4—6 ч после рождения ее содержание снижается до минимума (2,2—3,3 ммоль/л), оставаясь на таком уровне в течение последующих 3—4 дней. Повышенное потребление глюкозы тканями у новорожденных и период голодания после родов приводят к усилению гликогенолиза и использованию резервного гликогена и жира. Запас гликогена в печени у новорожденного в первые 6 ч жизни резко (примерно в 10 раз) сокращается, особенно при асфиксии и голодании. Содержание глюкозы в крови достигает возрастной нормы у доношенных новорожденных к 10—14-му дню жизни, а у недоношенных детей устанавливается лишь к 1—2-му месяцу жизни. В кишечнике новорожденных ферментативный гидролиз лактозы (основного углевода пищи в этот период) несколько снижен и увеличивается в грудном возрасте. Обмен галактозы у новорожденных интенсивнее, чем у взрослых.
Нарушения У. о. у детей при различных соматических заболеваниях носят вторичный характер и связаны с влиянием основного патологического процесса на этот вид обмена.
Лабильность механизмов регуляции углеводного и жирового обмена в раннем детском возрасте создает предпосылки для возникновения гипо- и гипергликемических состояний, ацетонемической рвоты. Так, например, нарушения У. о. при пневмонии у детей раннего возраста проявляются повышением в крови натощак концентраций глюкозы и лактата в зависимости от степени дыхательной недостаточности. Непереносимость углеводов выявляется при ожирении и обусловливается изменением секреции инсулина. У детей с кишечными синдромами часто выявляют нарушение расщепления и всасывания углеводов, при целиакии (см. Глютеновая болезнь) отмечают уплощение гликемической кривой после нагрузки крахмалом, дисахаридами и моносахаридами, а у детей раннею возраста с острыми энтероколитами и соледефицитным состоянием при обезвоживании наблюдают склонность к гипогликемии.В крови детей старшего возраста в норме отсутствуют галактоза, пентозы и дисахариды, у детей грудного возраста они могут появляться в крови после приема пищи, богатой этими углеводами, а также при генетически обусловленных аномалиях обмена соответствующих углеводов или углеводсодержащих соединений; в подавляющем большинстве случаев симптомы таких заболеваний проявляются у детей в раннем возрасте.
Для ранней диагностики наследственных и приобретенных нарушений У. о. у детей применяют этапную систему обследования с использованием генеалогического метода (см. Медицинская генетика), различных скрининг-тестов (см. Скрининг), а также углубленных биохимических исследований. На первом этапе обследования проводят определение в моче глюкозы, фруктозы, сахарозы, лактозы качественными и полуколичественными методами, проверяют значение рН кала. При получении результатов, заставляющих подозревать патологии) У. о., переходят ко второму этапу обследования: определению содержания глюкозы в моче и крови натощак количественными методами, построению гликемических и глюкозурических кривых, исследованию гликемических кривых после дифференцированных сахарных нагрузок, определению содержания глюкозы в крови после введения адреналина, глюкагона, лейцина, бутамида, кортизона, инсулина; в части случаев осуществляют прямое определение активности дисахаридаз в слизистой оболочки двенадцатиперстной и тонкой кишок и хроматографическую идентификацию углеводов крови и мочи. Для выявления нарушений переваривания и всасывания углеводов после установления значения рН кала определяют толерантность к моно- и дисахаридам с обязательным измерением содержания сахаров в кале и их хроматографической идентификацией до и после нагрузочных проб с углеводами При подозрении на энзимопатию (см. Ферментопатии) в крови и тканях определяют активность ферментов У. о., дефект синтеза (или снижение активности) которых подозревают клиницисты.
Для коррекции нарушенного У. о. при тенденции к гипергликемии применяют диетотерапию с ограничением жиров и углеводов. При необходимости назначают инсулин или другие гипогликемизирующие препараты; средства, способствующие повышению содержания глюкозы в крови, отменяют. При гипогликемии показана диета, богатая углеводами и белками.
Во время приступов гипогликемии вводят глюкозу, глюкагон, адреналин. При непереносимости отдельных углеводов назначают индивидуальную диету с исключением соответствующих сахаров из пищи больных. В случаях нарушений У. о., носящих вторичный характер, необходимо лечение основного заболевания.
Профилактика выраженных нарушений У. о. у детей заключается в их своевременном обнаружении. При вероятности наследственной патологии У. о. рекомендуется медико-генетическое консультирование. Выраженное неблагоприятное влияние декомпенсации сахарного диабета у беременных женщин на У. о. у плода и новорожденного диктует необходимость тщательной компенсации заболевания у матери на всем протяжении беременности и родов.
Библиогр.: Видершайн Г.Я. Биохимические основы гликозидозов, М., 1980; Гормональная регуляция функций детского организма в норме и патологии, под ред. М.Я. Студеникина и др., с. 33, М., 1978; Комаров Ф.И., Коровкин Б.Ф. и Меньшиков В.В. Биохимические исследования в клинике, с. 407, Л., 1981; Мецлер Д. Биохимия, пер. с англ., т. 2, М., 1980; Николаев А.Я. Биологическая химия, М., 1989; Розенфельд Е.Л. и Попова И.А. Врожденные нарушения обмена гликогена, М., 1989; Справочник по функциональной диагностике в педиатрии, под ред. Ю.Е. Вельтищева и Н.С. Кисляк, с. 107, М., 1979.
| Иллюстрации по статье: | ||||
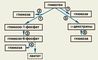 Рис. 2. Схема распада в о |  Рис. 1. Схема синтеза глю |
|||

Ваш комментарий