Онкология-
Опухолевые супрессоры и мутаторные гены
Б.П.Копнин
Российский онкологический научный центр им. Н.Н.Блохина РАМН, Москва
источник RosOncoWeb.Ru
01 02 03 04 05 06 073.4. Продукты гена INK4a - p16INK4a и pARF - регулируют активность pRb и р53
Следующим после р53 по частоте изменений в различных новообразованиях человека является ген INK4a, расположенный в коротком плече хромосомы 9 (сегмент 9р21). Его исключительной особенностью является одновременное кодирование двух негомологичных ядерных белков - p16INK4a и pARF (продукты альтернативных рамок считывания), каждый из которых выполняет супрессорные функции (Рис. 5).
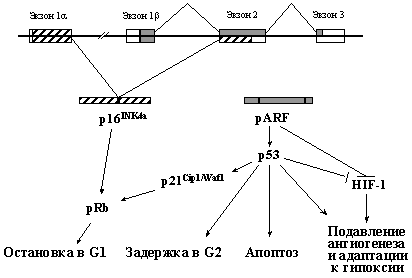
Рис. 5. Ген INK4a кодирует два негомологичных белка, выполняющих разные функции.
p16INK4a связывает циклин-зависимые киназы Cdk4 и Cdk6 и препятствует образованию их функционально активных комплексов с циклинами D, которые, фосфорилируя pRb, инициируют вход в S фазу клеточного цикла. pARF обладает способностью стабилизировать и активировать белок р53, нарушая его взаимодействие с белком Mdm2 (см. раздел 3.3). Кроме того, у него выявлены и р53-независимые функции. Так, он может также непосредственно взаимодействовать с основной мишенью pRb - белком E2F и блокировать его активность. Кроме того, он понижает стабильность транскрипционного фактора HIF-1 (см. 3.3.2 и 3.5.3), связывая его a-субъединицу.
Активность p16INK4a и pARF повышается при экспрессии ряда вирусных (E1A) или клеточных (RAS, RAF, MYC и др.) онкогенов. Такой эффект обусловлен присутствием в гене INK4a респонсивных элементов для транскрипционного фактора E2F, активируемого многими онкогенами. Таким образом, нормальное функционирование продуктов гена INK4a эффективно предотвращает дальнейшее размножение клеток, в которых произошла активация какого-либо из представителей большой группы онкогенов. Кроме того, экспрессия p16INK4a повышается при образовании межклеточных контактов, что обеспечивает контактное торможение размножения клеток.
У трансгенных мышей с гомозиготной нокаутом гена INK4a, вызывающим потерю экспрессии обоих его белковых продуктов, наблюдается высокая частота возникновения в молодом возрасте различных новообразований, преимущественно фибросарком и лимфом (также как у мышей с инактивированным р53). Интересно, что практически такая же картина наблюдается и у мышей с перестройкой гена INK4a, приводящей к потере экспрессии только белка pARF. В отличие от этого у мышей, утративших экспрессию p16INK4a, но сохранивших экспрессию pARF, нет существенного повышения частоты возникновения опухолей (отмечается лишь редкое возникновение меланом, не наблюдаемое в родительских линиях мышей). В культурах in vitro мышиные фибробласты, не экспессирующие оба продукта гена INK4a, как и фибробласты, в которых инактивирован только pARF, легко трансформируются онкогенами семейства RAS. В то же время для Ras-индуцированной трансформации клеток, не экспрессирующих только p16INK4a, необходимо дополнительное действие кооперирующих онкогенов, таких как MYC или E1, подобно тому, как это наблюдается в случае нормальных клеток.
У людей герминальные мутации в одном из двух аллелей гена INK4a ассоциированы с наследственной предрасположенностью к развитию меланом кожи (синдром диспластических невусов). Часть из этих мутаций инактивирует только p16INK4a, не нарушая функцию pARF. В связи с этим предполагается, что синдром диспластических невусов связан с нарушением функции именно p16INK4a. В клетках наследственных и спорадических меланом обнаруживаются изменения обоих аллелей гена INK4a, т.е. данный ген ведет себя как классический опухолевый супрессор. Чаще всего вторая (соматическая) мутация представляет собой делецию гена INK4a, ведущую к инактивации как p16INK4a, так и pARF. Мутации, делеции и метилирование гена INK4a, вызывающие потерю экспрессии одного или обоих его белковых продуктов, часто наблюдаются не только в наследственных и спорадических меланомах, но и в большой группе других ненаследственных новообразований: раке поджелудочной железы, пищевода, желчных путей, мочевого пузыря, Т- и В-клеточных острых лимфолейкозах, мезотелиомах, анапластических астроцитомах, глиобластомах и др.
3.5. Опухолевый супрессор PTEN регулирует клеточный цикл и апоптоз, модулируя сигнальный путь PI3K-PKB/Akt
К опухолевым супрессорам, часто поражаемым в различных новообразованиях человека, относится и ген PTEN. Его герминальные мутации вызывают болезнь Коудена, заключающуюся в наследственном предрасположении к развитию гамартом, чаще всего в мозге, молочной и щитовидной железах. Инактивация PTEN характеризует также и значительную часть различных ненаследственных опухолей - глиомы, менингиомы, меланомы, раки почки, матки, молочной и предстательной желез. При этом на начальных стадиях заболевания выявляется, как правило, делеция только одного из аллелей гена PTEN, тогда как в опухолях, находящихся на поздних стадиях развития, чаще инактивированы оба аллеля. У трансгенных мышей с нокаутом одного аллеля гена PTEN отмечается развитие в молодом возрасте самых разных новообразований: аденокарцином кишечника, лейкозов, лимфосарком, герминальных опухолей и др. Белковый продукт гена PTEN состоит из 403 аминокислот и имеет высокую степень гомологии с фосфатазами двойной специфичности (дефосфорилируют и тирозиновые, и серин/треониновые аминокислотные остатки). Его особенностью является способность дефосфорилировать не только белки, но и липиды, в частности фосфатидил-инозит(3,4,5)трифосфат (PIP-3) - важнейшую мишень фосфатидил-инозитид-3'-киназы (PI3K), участвующую в регуляции клеточного цикла и апоптоза. Именно эта активность белка PTEN ответственна, по-видимому, за его супрессорную функцию. Об этом свидетельствует в частности тот факт, что большинство мутаций, обнаруживаемых в опухолевых клетках, картируются в фосфатазном домене и отменяют дефосфорилирование PIP-3, ингибируя таким образом сигнальный путь PI3K-PKB/Akt. Так как онкоген PKB/Akt подавляет митохондриальный путь индукции апоптоза по нескольким механизмам (ингибирует Bad, активирует Bcl2 и т.д.), потеря функции PTEN делает клетки менее чувствительными ко многим апоптогенным стимулам. В клетках с инактивированным PTEN наблюдается также и стимуляция клеточной пролиферации (повышение экспрессии PKB/Akt увеличивает уровень циклина D). Восстановление функции PTEN в опухолевых клетках приводит, в зависимости от клеточного контекста, либо к апоптозу (рак простаты), либо к остановке клеточного цикла в G1 (глиобластома, рак почки). Следует заметить, что клетки глтобластомы и рака почки становятся при этом сенсибилизированными к действию дополнительных апоптогенных стимулов. PTEN участвует, по-видимому, и в регуляции адгезии и миграции клеток. Его N-концевой домен гомологичен тензину - белку, взаимодействующему в фокальных контактах с актиновым цитоскелетом. Дефосфорилируя тирозиновые остатки киназы фокальных контактов FAK, PTEN может ингибировать образование фокальных контактов и распластывание клеток, а также их движение.